baimsizdir. Günümüze kadar yedi GRK tes-
pit edilmitir.
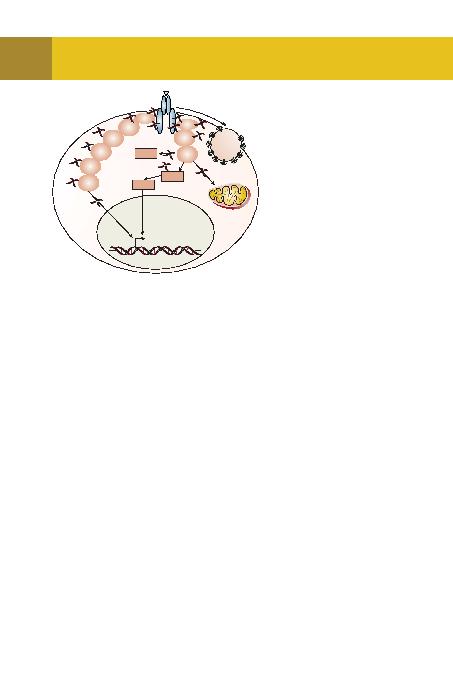
ini ayrilir ve reseptörün sitoplazmaya uzanan
kuyruu yanisira bazi intrasellüler domainleri,
GRK tarafindan fosfatlanir. Bu sayede beta-
arrestinler aktive olur. Bu moleküller reseptö-
rün G proteinlerle olan irtibatini keser. Bunun
yanisira, ligandina balanmi ve sinyal üre-
timine hazir GPCR'leri endositoza uratarak
hücreden izole eder. Bu iki mekanizma sayesin-
de hormon bali GPCR'lerin hücre içine daha
fazla uyari göndermesi engellenir.
sensitizasyon fenomeni olarak adlandirilir.
lerinden baimsiz bir sinyal ilevine sahip ol-
duu da bilinmektedir.
olarak 60 reseptör bulunur. Reseptörlerin bü-
yük bir bölümünde ligand-balayici ekstrasel-
lüler domain ve intrasellüler tirozin-kinaz do-
mainden oluan yapi görülür. Epidermal büyü-
me faktörü (EBF), fibroblast büyüme faktörü
birçok büyüme faktörünün reseptörü ise sade-
ce transmembran polipepetidlerinden oluur.
Polipeptidler, ligandin balanmasindan sonra
dipeptidlere veya oligopeptidlere parçalanir.
törleri ise birbirine disülfit balarla balanmi
kovalent dimerlerden oluur.
lanmasi otofosforilasyon ilemini balatir.
IGF-2 reseptörü mannoz-6-fosfat içeren ve
hücre zarinda tek baina bulunan büyük bir
transmembran reseptördür. Bu reseptörün
görevi balayici proteinden IGF-II'yi ayirmak-
tir.
ki aminoasit dizilimi bakimindan %84'lük bir
oranla, insülin reseptörüne benzer bir yapida-
dir.
iki izoformdan oluur (A ve B). zoform yapilar
INSR geninin 11. ekzonundaki farkliliktan kay-
naklanir.
finite ile balanirlar. Reseptör ve hormonlarin
yapisal benzerliinden dolayi çapraz balanma
bir affinite ile balanirken, IGF-1 ve insülin B
reseptörü öznel olmayan moleküle düük bir
affinite ile balanir.
septörleri ilevsel bir hibrid reseptör olutur-
mak üzere birleebilirler. Bu sayede insülin
ve IGF'ler çapraz bir etki için daha büyük bir
potansiyele sahip olur. nsülin-IGF-reseptör
sisteminin özgünlüünü belirleyen unsurlar;
Hormonlarin biyoyararlanimi, reseptörün
ekspresyonu (insülin reseptörü izoformlarinin
ekspresyonu dokuya ve organizmanin geliim
evresine göre deiir), balanma affinitesinde-
ki farklilik ve iletim yolaini aktive edebilme
becerisidir.
reseptör bulunur. Büyüme hormonu reseptörü
(BHR), prolaktin reseptörü (PRLR) ve leptin
reseptörü (LR) bunlardan bazilaridir. Bu resep-
törler ilikide olduklari kinazlari aktive ederler,
ancak intrensek katalitik aktivasyon kaskadina
katilmazlar. Reseptörler hem kisa hem de uzun
formlarda oluur. Büyüme hormonu BHR'ye
balandii zaman, reseptörün ekstrasellüler
domaini parçalanarak dolaima geçer. Suda
eriyebilen bu bölüm BH-balayici protein'i
(BHBP) oluturur.
bir reseptördür.
nindeki farkliliktan kaynaklanir.
proteinlerinden oluur. Bu proteinler, tirozin
kinaz Janus kinaz 2 (JAK2) adli enzimin ile-
vinde rol alir. Bu enzim
ilev gören "sinyal iletimi ve transkripsiyon
aktivatörleri"ni (STAT) aktive eder ve gen
traskripsiyon ilemini balatir.
sinyal iletim yolaklari da aktive edilir. Örne-
in, MAP kinaz, PI3K/Akt, Ras-benzeri küçük
GTP'azlar ve dolayli yoldan ikincil haberciler
BH, prolaktin ve leptin reseptörleri tarafindan
aktive edilirken,
(CIS), sitokin reseptör ailesi için negatif bir
feedback oluturur. BH, prolaktin ve leptin
reseptörünün uyarilmasi ile balayan süreç
birbirinden farkli SSS/SIS proteinini de aktif
hale getirir. Böylece STAT sinyali inhibe edile-
rek, hormonun etkisi sonlandirilir.
üzerinden sinyal iletimi balatirlar. Onlar bu
SOS
subünitlerinin sitoplazma içine doöru uzanan uçlar¿nda intrensek tirozin kinaz aktivitesi vard¿r. Hormo-
nun reseptöre baölanmas¿ otofosforilasyon iùlevini aktive eder. Fosfatlanan tirozin molekülleri, adaptör
moleküllerin reseptöre baölanmas¿n¿ saölar. Adaptör moleküller; SRC homolog 2 domain-içeren protein
(SHC) ve insülin reseptör substrat 1 (IRS1) ailesidir. Bu moleküller MAP kinaz ve PI3K kaskad¿n¿ fosfat-
lar ve aktive ederler. Bu kaskatta yer alan ekstrasellüler-regüle edilen protein kinaz (ERK), Akt (Protein
kinaz B) ve rapamisinin memelilerdeki hedefi (RMH) aktive olur. Sonuç olarak, tirozin-kinaz reseptörleri,
hücrede bölünme ve metabolik aktivitelerin yan¿s¿ra yaùamsal fonksiyonlar¿n devam¿ için mitokondri ve
nükleusa (NF-kappa-B translokasyon; c-myc ve c-Fos'un indüksiyonu gibi) yönelik sinyal iletiminde rol
oynarlar.