mesine kari dirençlidirler. Bunlar aktive mak-
rofajlarca öldürülürler: stirahat durumunda
iken makrofajlarin patojenleri öldürme kabi-
liyetleri sinirli ise de -interferon ve TNF-
ile aktive olmalari intraselüler öldürmelerini
dramatik olarak iyiletirir (
daha aktif hale gelebilir, sonra agrege olur ve
granülom olutururlar.
moleküllerini eksprese etmeleri ile ayiredilen
bir lenfosit alt grubudur. Hedef hücreleri par-
çalayan sitotoksik mekanizmalari kullanarak
virüslerle enfekte hücreyi ve tümör hücresini
seçici olarak tanir ve öldürürler. Hedef hücre ile
temasi takiben organeller kalsiyum-baimli bir
mekanizma ile hedefe yönelirler. Bunu takiben,
granüllerden granzim A, perforin ve TGF- or-
tama salinir. NK hücreleri yüzeylerindeki CD16
(FcRIII) ve CD56 (NCAM-1) ekspresyonlari
ile taninir ve dolaimdaki lenfositlerin %10-
15'ini olutururlar. Matür NK hücreleri dalak,
akcier ve karacierde bulunabilirler. Gebeliin
birinci trimesteri sirasinda plasental desiduada
da bulunurlar. NK hücreleri sitolitik fonksiyon-
larini düzenlemede bir dizi uyarici ve kisitlayici
reseptörü kullanirlar. NK hücrelerinde grup 2 D
üyesi bir reseptörün viral enfeksiyon veya ma-
lign dönüüm gibi stres durumlarinda hücrele-
re eksprese edilen molekülleri tanima ve aktive
edici fonksiyonlari vardir.
deitirmede önemli bir görevi vardir. Kalitsal
ve akkiz NK hücre defektleri hemofagositik len-
durumla ilgilidir (Bölüm 463'e bakiniz).
olan antimikrobiyal peptidleri, C reaktif pro-
tein ve mannoz balayici lektin gibi serum
proteinlerini içerir. Kompleman sistemi Bölüm
189'da daha ayrintili tartiilacaktir.
doal immün sistem hücrelerinin birçok mik-
roba ait ortak moleküler motiflerini tanima-
sinin tersine, adaptif immün cevaplar antijen
adi verilen benzersiz peptidlere kari geliir.
Bu özellik immün spesifisite olarak adlandiri-
lir.
yi salar. Ayni antijenle daha sonra tekrardan
maruziyet, daha hizli ve güçlü bir immün ce-
vapla sonuçlanir ve buna immünolojik hafiza
denir. Adaptif immün cevaplar, sirasiyla T ve B
hücrelerinin aracilik ettii hücresel ve hümoral
immün cevaplara ayrilir.
T-hücre reseptörü bir zinciri ve bir zinci-
ri bulundururken, yaklaik %5'i bir zinciri
ve bir zinciri içerir. Zincirlerin her biri hem
sabit hem deiken gen segmentlerine sahiptir.
Deiken bölgeler, gen segmentlerinin parça
parça birleerek kaynamasi ile DNA'nin ye-
niden düzenlenmesi ve tam olarak bilinmeyen
baka çeitlilik mekanizmalari yoluyla oluur.
Birbirinden farkli 10
sayida farkli reseptör bazilarinin enfeksiyon
ajanlarindan çok kendi proteinlerini balaya-
bilmesinin mümkün olduunu düündürmek-
tedir.
IL-8
P Selektin
E Selektin
PSGL 1
PECAM
MIC-2
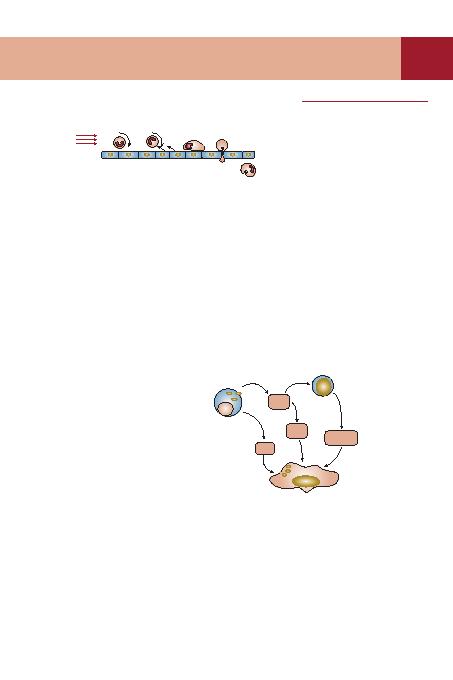
n¿zca itici ve çekici kuvvetlerin etkisi alt¿nda selektin ailesi ile etkileùir. Endotel hücresi üzerinde selek-
tin protein ailesi inflamatuvar arac¿ moleküllerin etkisiyle say¿ca artt¿r¿l¿r. Temas sonras¿nda endotel
hücresince trombosit aktive edici faktör (PAF) ve IL-8'in üretimi, nötrofil
içinden gerçekleùebilir. Endotelyal bariyerden hücrenin geçiùine ek moleküller rehberlik eder. Nötrofil,
doku aral¿ö¿ndan kemotaktik gradiente uygun olarak bakteriye doöru göç eder. Oluùacak fagositozda
mannoz reseptörlerine, scavenger (çöpçü) reseptörlerine, Fc reseptörlere ve kompleman reseptörlerine
baölanma rol oynar. Bakteri, içeri al¿nma sonras¿nda birçok antimikrobiyal maddeye maruz kal¿r. Sonuç-
ta sürecin haddinden fazla uzamamas¿ için nötrofil fagositozdan sonra ölür. ICAM, hücre-aras¿ adhezyon
molekülleri; IL-8, interlökin 8; MIC-2, makrofaj inhibe edici sitokin-2; PAF, trombosit aktive edici faktör;
PECAM, trombosit/endotel hücre adhezyon molekül 1; psgl-1, p-selektin glikoprotein baölay¿c¿ ligand-1.
(
içi patojene karù¿ as¿l savunmad¿r. Makrofajlar nötrofiller ile ayn¿ mikrobisidal ürünleri içermez ve bir
öldürme program¿n¿ aktive etmelidirler. Makrofajlar¿n aktivasyonu ve hücre-içi öldürmeleri için hem
veya patern-tan¿ma reseptörleri taù¿yan ve NF-
lenfositlerin
T hücre bileùenini etkileyen HIV gibi hastal¿klar veya sitokin düzenini bozan TNF-