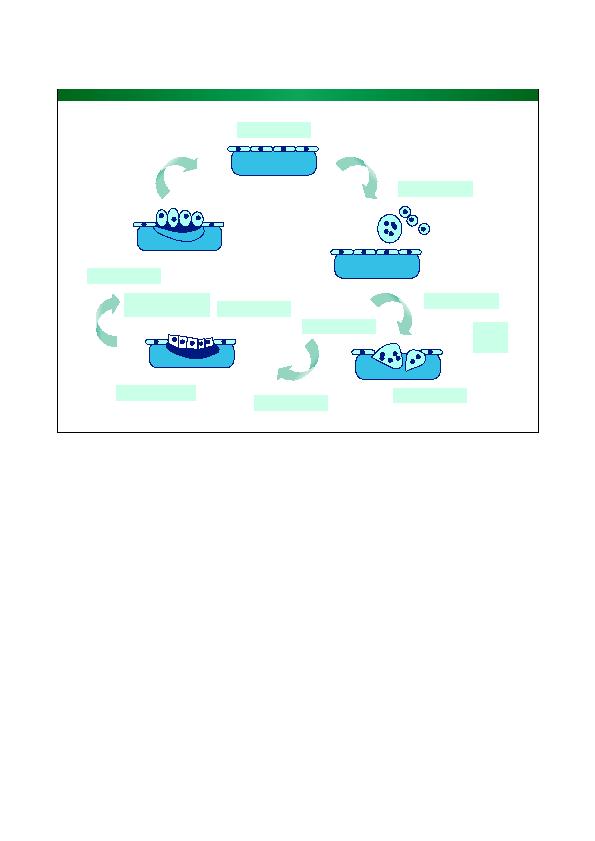
skelet aan overbelasting en herhaaldelijke trauma's. Remo
dellering is afhankelijk van de werking van gespecialiseer-
de cellen: enerzijds botresorptie door de osteoclasten, die
verouderd bot elimineren, dat niet langer is aangepast aan
de mechanische weerstand en/of de metabole functies en
anderzijds botvorming door de osteoblasten. Oude osteo
blasten laten zichzelf `begraven' in het bot en zorgen voor
de mechanotransductie door op belasting van het skelet
te reageren met de secretie van sclerostine. Dit cytokine
circuleert in de onderling verbonden kanaaltjes die com
municeren met het oppervlak. Dit proces herlanceert de
remodellering door stimulatie van het RANKL-RANK-
OPG-systeem in de meercellige functionele basiseenheden
(BMU's) waarvan er in de botten miljoenen aanwezig zijn
(1). Het normale remodelleringsritme is moeilijk te bepa
len, zelfs bij vrouwen voor de menopauze, het referentie
punt. De serumconcentraties van Cterminaal telopeptide
van type 1collageen (CTX), een katabool residu van de
resorptie, en het Nterminaal propeptide van procollageen
type 1 (P1NP) (resultaat van de botvorming) zijn hoger in
Frankrijk dan in het Verenigd Koninkrijk... Ongeveer 2,5
procent van de normale jonge vrouwen voor hun meno
pauze heeft concentraties van deze markers die lager zijn
dan de onderste grenswaarden van de laboratoriumrefe
rentiewaarden (2).
volledige remodelleringcyclus (resorptie rustfase vorming
rustfase) 8,2 maanden (afwijkingen 3,4-54 maanden) (3).
Er is een koppeling in de cellulaire acties van de BMU's, die
20 keer zo talrijk zijn in het trabeculaire bot dan in het cor
ticale bot. Het corticale bot vertegenwoordigt 80 procent
van de botmassa (Figuur 2).
het botmetabolisme in het trabeculaire bot dan in het cor
ticale bot in situaties met een hoge remodellering (voor
beeld: de menopauze). Het botverlies dat in deze situaties
wordt vastgesteld, kan worden verklaard door een nega
tief evenwicht tussen de osteoclastische en de osteoblas
tische activiteit, die beide versnellen. Maar de osteoblasten
hebben moeite om de beweging te volgen in de remodel
leringseenheden waarvan de activeringsfrequentie is ver
sneld. Tegenwoordig kan het globale botremodellerings
niveau worden berekend aan de hand van een meting van
uiteenlopende biologische markers. Merk wel op dat het
nog niet mogelijk is om biologisch het onderscheid te ma
ken tussen remodellering van het trabeculaire bot en remo
dellering van het corticale gedeelte: het is ook niet moge
lijk om bij kinderen in de groei het onderscheid te maken
tussen de botmodelleringswaarde (die de vorm van het bot
aanpast in functie van de verlenging ervan) (Figuur 3), en