- Page 1
- Page 2
- Page 3
- Page 4
- Page 5
- Page 6
- Page 7
- Page 8
- Page 9
- Page 10
- Page 11
- Page 12
- Page 13
- Page 14
- Page 15
- Page 16
- Page 17
- Page 18
- Page 19
- Page 20
- Page 21
- Page 22
- Page 23
- Page 24
- Page 25
- Page 26
- Page 27
- Page 28
- Page 29
- Page 30
- Page 31
- Page 32
- Page 33
- Page 34
- Page 35
- Page 36
- Page 37
- Page 38
- Page 39
- Page 40
- Page 41
- Page 42
- Page 43
- Page 44
- Page 45
- Page 46
- Page 47
- Page 48
- Page 49
- Page 50
- Page 51
- Page 52
- Page 53
- Page 54
- Page 55
- Page 56
- Page 57
- Page 58
- Page 59
- Page 60
- Page 61
- Page 62
- Page 63
- Page 64
- Page 65
- Page 66
- Page 67
- Page 68
- Page 69
- Page 70
- Flash version
© UniFlip.com
- Page 2
- Page 3
- Page 4
- Page 5
- Page 6
- Page 7
- Page 8
- Page 9
- Page 10
- Page 11
- Page 12
- Page 13
- Page 14
- Page 15
- Page 16
- Page 17
- Page 18
- Page 19
- Page 20
- Page 21
- Page 22
- Page 23
- Page 24
- Page 25
- Page 26
- Page 27
- Page 28
- Page 29
- Page 30
- Page 31
- Page 32
- Page 33
- Page 34
- Page 35
- Page 36
- Page 37
- Page 38
- Page 39
- Page 40
- Page 41
- Page 42
- Page 43
- Page 44
- Page 45
- Page 46
- Page 47
- Page 48
- Page 49
- Page 50
- Page 51
- Page 52
- Page 53
- Page 54
- Page 55
- Page 56
- Page 57
- Page 58
- Page 59
- Page 60
- Page 61
- Page 62
- Page 63
- Page 64
- Page 65
- Page 66
- Page 67
- Page 68
- Page 69
- Page 70
- Flash version
© UniFlip.com
176
B Ö LÜ M 5
VİRUS
İmmün Sistem Hastalıkları
gp120 CD4'e bağlanır gp41 gp120 VİRÜSÜN GİRİŞİ
Yapısal değişiklik
gp41 membran gp120 ve CD4 CCR5'e bağlanır penetrasyonu Membrana füzyon
HIV membranının konak hücre membranı ile füzyonu; viral genomun sitoplazmaya girişi
Sitokin Sitokin reseptörü
CD4
Kemokin reseptörü
HIV RNA genomu Proviral DNA'nın ters transkriptaz-aracılı sentezi Hücrenin sitokin ile aktivasyonu; HIV genomunun transkripsiyonu; viral RNA'ların sitoplazmaya taşınması
VİRÜSÜN REPLİKASYONU HIV proteinlerinin sentezi; virion çekirdek yapısının oluşması HIV çekirdek yapısı
Provirüsün konak hücre genomuna entegrasyonu
HIV RNA kopyası
HIV DNA provirüs Nükleus
VİRÜSÜN SALINMASI Tomurcuklanma ve matür virionun salınması
Yeni HIV virionu
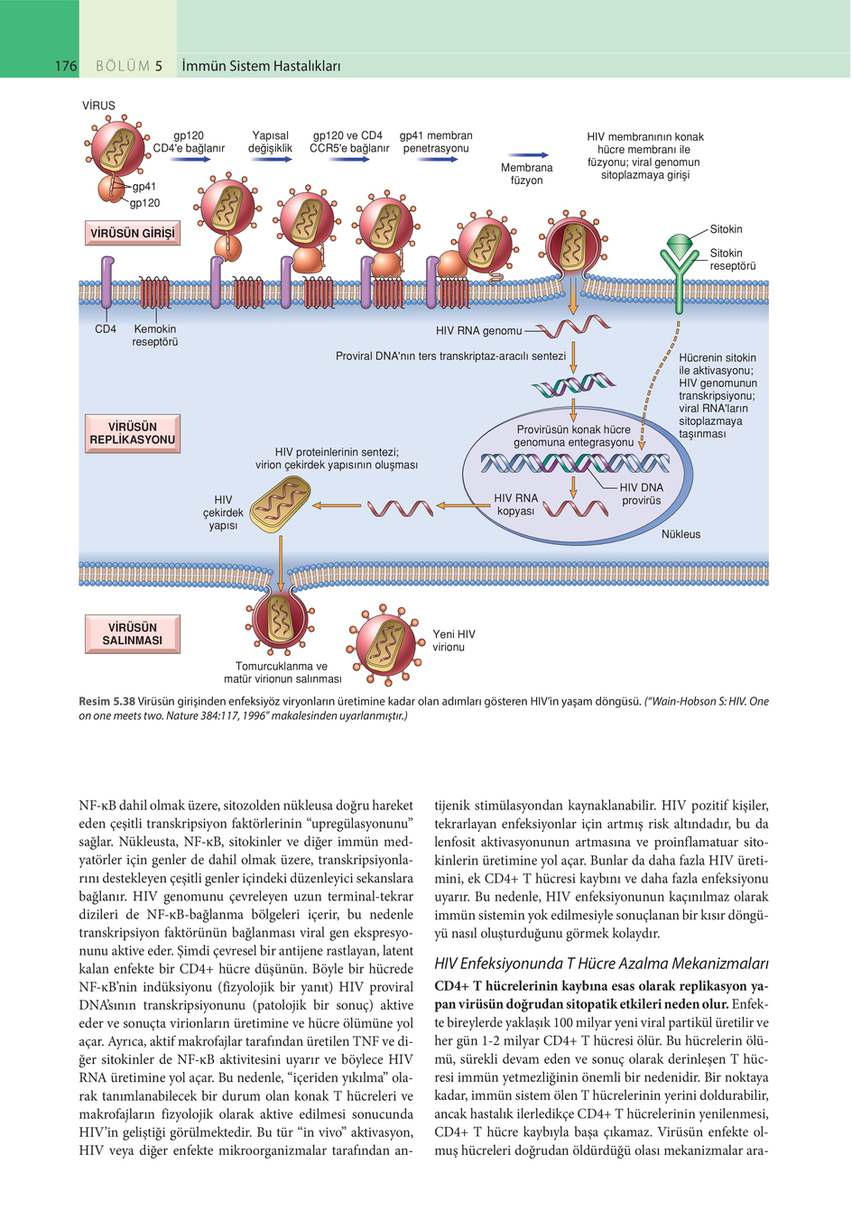
Resim 5.38 Virüsün girişinden enfeksiyöz viryonların üretimine kadar olan adımları gösteren HIV’in yaşam döngüsü. (“Wain-Hobson S: HIV. One on one meets two. Nature 384:117, 1996” makalesinden uyarlanmıştır.)
NF-κB dahil olmak üzere, sitozolden nükleusa doğru hareket eden çeşitli transkripsiyon faktörlerinin “upregülasyonunu” sağlar. Nükleusta, NF-κB, sitokinler ve diğer immün medyatörler için genler de dahil olmak üzere, transkripsiyonlarını destekleyen çeşitli genler içindeki düzenleyici sekanslara bağlanır. HIV genomunu çevreleyen uzun terminal-tekrar dizileri de NF-κB-bağlanma bölgeleri içerir, bu nedenle transkripsiyon faktörünün bağlanması viral gen ekspresyonunu aktive eder. Şimdi çevresel bir antijene rastlayan, latent kalan enfekte bir CD4+ hücre düşünün. Böyle bir hücrede NF-κB’nin indüksiyonu (fizyolojik bir yanıt) HIV proviral DNA’sının transkripsiyonunu (patolojik bir sonuç) aktive eder ve sonuçta virionların üretimine ve hücre ölümüne yol açar. Ayrıca, aktif makrofajlar tarafından üretilen TNF ve diğer sitokinler de NF-κB aktivitesini uyarır ve böylece HIV RNA üretimine yol açar. Bu nedenle, “içeriden yıkılma” olarak tanımlanabilecek bir durum olan konak T hücreleri ve makrofajların fizyolojik olarak aktive edilmesi sonucunda HIV’in geliştiği görülmektedir. Bu tür “in vivo” aktivasyon, HIV veya diğer enfekte mikroorganizmalar tarafından an-
tijenik stimülasyondan kaynaklanabilir. HIV pozitif kişiler, tekrarlayan enfeksiyonlar için artmış risk altındadır, bu da lenfosit aktivasyonunun artmasına ve proinflamatuar sitokinlerin üretimine yol açar. Bunlar da daha fazla HIV üretimini, ek CD4+ T hücresi kaybını ve daha fazla enfeksiyonu uyarır. Bu nedenle, HIV enfeksiyonunun kaçınılmaz olarak immün sistemin yok edilmesiyle sonuçlanan bir kısır döngüyü nasıl oluşturduğunu görmek kolaydır.
HIV Enfeksiyonunda T Hücre Azalma Mekanizmaları
CD4+ T hücrelerinin kaybına esas olarak replikasyon yapan virüsün doğrudan sitopatik etkileri neden olur. Enfekte bireylerde yaklaşık 100 milyar yeni viral partikül üretilir ve her gün 1-2 milyar CD4+ T hücresi ölür. Bu hücrelerin ölümü, sürekli devam eden ve sonuç olarak derinleşen T hücresi immün yetmezliğinin önemli bir nedenidir. Bir noktaya kadar, immün sistem ölen T hücrelerinin yerini doldurabilir, ancak hastalık ilerledikçe CD4+ T hücrelerinin yenilenmesi, CD4+ T hücre kaybıyla başa çıkamaz. Virüsün enfekte olmuş hücreleri doğrudan öldürdüğü olası mekanizmalar ara-