- Page 1
- Page 2
- Page 3
- Page 4
- Page 5
- Page 6
- Page 7
- Page 8
- Page 9
- Page 10
- Page 11
- Page 12
- Page 13
- Page 14
- Page 15
- Page 16
- Page 17
- Page 18
- Page 19
- Page 20
- Page 21
- Page 22
- Page 23
- Page 24
- Page 25
- Page 26
- Page 27
- Page 28
- Page 29
- Page 30
- Page 31
- Page 32
- Page 33
- Page 34
- Page 35
- Page 36
- Page 37
- Page 38
- Page 39
- Page 40
- Page 41
- Page 42
- Page 43
- Page 44
- Page 45
- Page 46
- Page 47
- Page 48
- Page 49
- Page 50
- Page 51
- Page 52
- Page 53
- Page 54
- Page 55
- Page 56
- Page 57
- Page 58
- Page 59
- Page 60
- Page 61
- Page 62
- Page 63
- Page 64
- Page 65
- Page 66
- Page 67
- Page 68
- Page 69
- Page 70
- Flash version
© UniFlip.com
- Page 2
- Page 3
- Page 4
- Page 5
- Page 6
- Page 7
- Page 8
- Page 9
- Page 10
- Page 11
- Page 12
- Page 13
- Page 14
- Page 15
- Page 16
- Page 17
- Page 18
- Page 19
- Page 20
- Page 21
- Page 22
- Page 23
- Page 24
- Page 25
- Page 26
- Page 27
- Page 28
- Page 29
- Page 30
- Page 31
- Page 32
- Page 33
- Page 34
- Page 35
- Page 36
- Page 37
- Page 38
- Page 39
- Page 40
- Page 41
- Page 42
- Page 43
- Page 44
- Page 45
- Page 46
- Page 47
- Page 48
- Page 49
- Page 50
- Page 51
- Page 52
- Page 53
- Page 54
- Page 55
- Page 56
- Page 57
- Page 58
- Page 59
- Page 60
- Page 61
- Page 62
- Page 63
- Page 64
- Page 65
- Page 66
- Page 67
- Page 68
- Page 69
- Page 70
- Flash version
© UniFlip.com
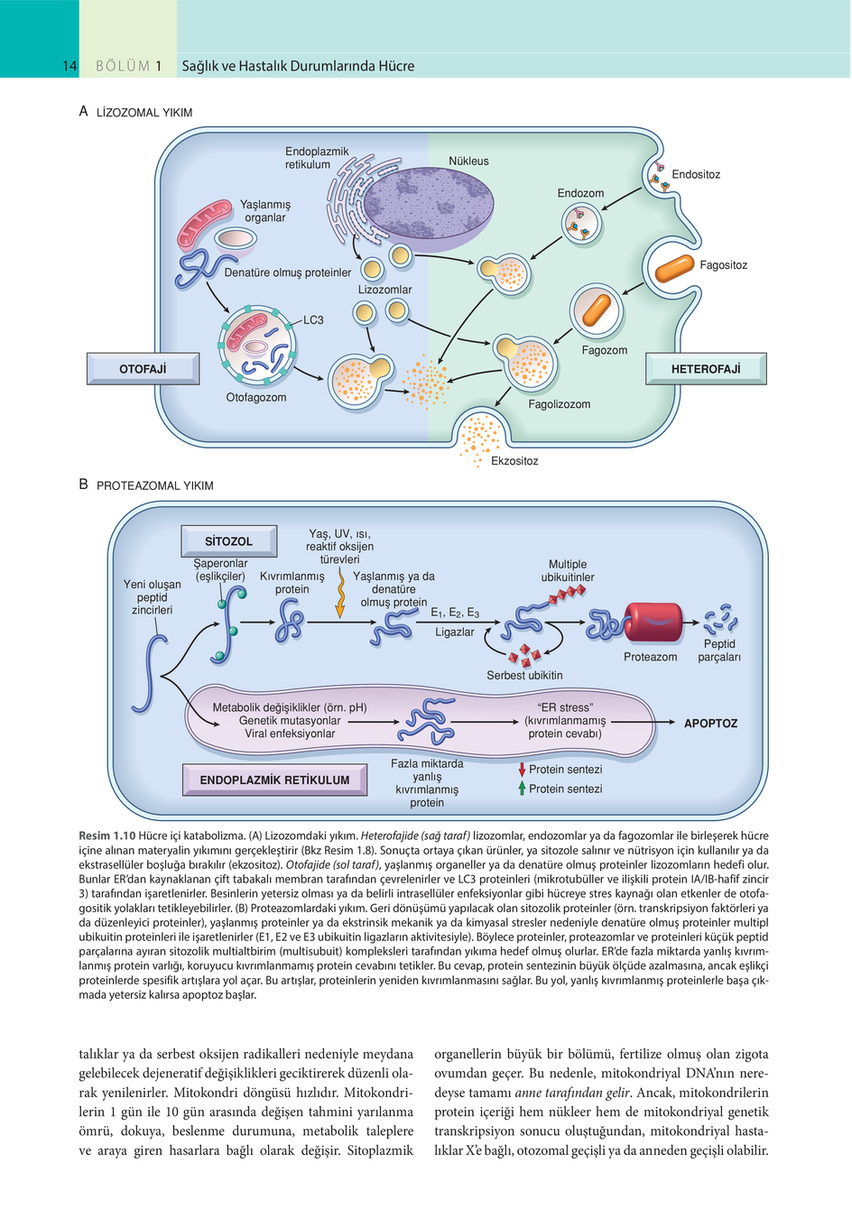
14
A
B Ö LÜ M 1
Sağlık ve Hastalık Durumlarında Hücre
LİZOZOMAL YIKIM Endoplazmik retikulum
Nükleus Endositoz Endozom
Yaşlanmış organlar
Denatüre olmuş proteinler Lizozomlar LC3 Fagozom OTOFAJİ Otofagozom Fagolizozom
Fagositoz
HETEROFAJİ
Ekzositoz
B
PROTEAZOMAL YIKIM
SİTOZOL Şaperonlar (eşlikçiler)
Yeni oluşan peptid zincirleri
Yaş, UV, ısı, reaktif oksijen türevleri Kıvrımlanmış Yaşlanmış ya da protein denatüre olmuş protein E1, E2, E3 Ligazlar
Multiple ubikuitinler
Proteazom Serbest ubikitin Metabolik değişiklikler (örn. pH) Genetik mutasyonlar Viral enfeksiyonlar Fazla miktarda yanlış kıvrımlanmış protein “ER stress” (kıvrımlanmamış protein cevabı) Protein sentezi Protein sentezi
Peptid parçaları
APOPTOZ
ENDOPLAZMİK RETİKULUM
Resim 1.10 Hücre içi katabolizma. (A) Lizozomdaki yıkım. Heterofajide (sağ taraf) lizozomlar, endozomlar ya da fagozomlar ile birleşerek hücre içine alınan materyalin yıkımını gerçekleştirir (Bkz Resim 1.8). Sonuçta ortaya çıkan ürünler, ya sitozole salınır ve nütrisyon için kullanılır ya da ekstrasellüler boşluğa bırakılır (ekzositoz). Otofajide (sol taraf), yaşlanmış organeller ya da denatüre olmuş proteinler lizozomların hedefi olur. Bunlar ER’dan kaynaklanan çift tabakalı membran tarafından çevrelenirler ve LC3 proteinleri (mikrotubüller ve ilişkili protein IA/IB-hafif zincir 3) tarafından işaretlenirler. Besinlerin yetersiz olması ya da belirli intrasellüler enfeksiyonlar gibi hücreye stres kaynağı olan etkenler de otofagositik yolakları tetikleyebilirler. (B) Proteazomlardaki yıkım. Geri dönüşümü yapılacak olan sitozolik proteinler (örn. transkripsiyon faktörleri ya da düzenleyici proteinler), yaşlanmış proteinler ya da ekstrinsik mekanik ya da kimyasal stresler nedeniyle denatüre olmuş proteinler multipl ubikuitin proteinleri ile işaretlenirler (E1, E2 ve E3 ubikuitin ligazların aktivitesiyle). Böylece proteinler, proteazomlar ve proteinleri küçük peptid parçalarına ayıran sitozolik multialtbirim (multisubuit) kompleksleri tarafından yıkıma hedef olmuş olurlar. ER’de fazla miktarda yanlış kıvrımlanmış protein varlığı, koruyucu kıvrımlanmamış protein cevabını tetikler. Bu cevap, protein sentezinin büyük ölçüde azalmasına, ancak eşlikçi proteinlerde spesifik artışlara yol açar. Bu artışlar, proteinlerin yeniden kıvrımlanmasını sağlar. Bu yol, yanlış kıvrımlanmış proteinlerle başa çıkmada yetersiz kalırsa apoptoz başlar.
talıklar ya da serbest oksijen radikalleri nedeniyle meydana gelebilecek dejeneratif değişiklikleri geciktirerek düzenli olarak yenilenirler. Mitokondri döngüsü hızlıdır. Mitokondrilerin 1 gün ile 10 gün arasında değişen tahmini yarılanma ömrü, dokuya, beslenme durumuna, metabolik taleplere ve araya giren hasarlara bağlı olarak değişir. Sitoplazmik
organellerin büyük bir bölümü, fertilize olmuş olan zigota ovumdan geçer. Bu nedenle, mitokondriyal DNA’nın neredeyse tamamı anne tarafından gelir. Ancak, mitokondrilerin protein içeriği hem nükleer hem de mitokondriyal genetik transkripsiyon sonucu oluştuğundan, mitokondriyal hastalıklar X’e bağlı, otozomal geçişli ya da anneden geçişli olabilir.