gnétisation nette se déplace de l'axe Z
longitudinal vers le plan XY transversal.
En outre, l'onde entraîne la rotation en
phase des noyaux dans le plan XY (os-
cillations cohérentes). Lorsque les noyaux
sont en déphasage, cela provoque un
signal magnétique détectable, qui est
enregistré. Étant donné que les protons
préfèrent un état d'énergie faible, ils
cèderont l'énergie qu'ils ont captée et re-
tourneront à un état d'équilibre en se réali-
gnant avec l'axe Z longitudinal (champ
magnétique; B0). Ce processus porte le
nom de relaxation et peut se subdiviser en
deux paramètres indépendants: la relaxa-
tion T1 et T2 (Figure 1).
nale, indique le degré de rétablissement
de la composante longitudinale du vec-
teur de magnétisation, et est défini
comme le temps (en millisecondes) né-
cessaire pour que la magnétisation lon-
gitudinale atteigne 63% de sa valeur
finale (17). Cette composante du signal de
RM reflète les aspects structurels et est
tions de l'état du muscle.
T2, ou temps de relaxation transver-
sale, reflète le degré de déclin du vec-
teur de magnétisation dans le plan
transversal ou XY. T2 est défini comme
le temps (en millisecondes) nécessaire
pour que le signal transversal atteigne
37% (1 divisé par la constante e) de sa
valeur initiale (17). Contrairement à
T1, T2 est sensible aux modifications
du temps de relaxation de l'eau dans le
muscle.
l'IRM-fm
allongement du temps de relaxation T2
de l'eau dans le muscle après un exer-
cice. De manière spécifique, l'activité
entraîne un déclin plus lent du signal de
l'eau dans le muscle. Par rapport à l'ima-
gerie des muscles à l'état de repos, ceci
provoque une amélioration de l'intensité
du signal des muscles activés, avec en
conséquence une image plus nette et
plus claire des muscles activés sur les
(15).
Plusieurs études ont déjà été réalisées
pour expliquer le mécanisme physiolo-
gique sous-jacent de ce glissement du
temps de relaxation T2 (18-20). L'expli-
cation la plus simple est que le courant
de liquide entrant durant l'activité est
associé à une accumulation d'osmolytes
(phosphate, lactate, sodium) dans le cy-
toplasme et que leur présence allonge le
temps de relaxation de l'eau dans le
muscle (15). La quantité totale d'eau
dans le muscle est constituée de plu-
sieurs composantes, telles que 1) l'eau
intracellulaire liée aux protéines (34%),
2) l'eau intracellulaire libre (49%) et 3)
l'eau extracellulaire (14%), chaque com-
posante présentant une modification de
son temps de relaxation T2 respectif (21).
La somme des effets des modifications
de ces composantes entraîne l'élévation
nette de T2, induite par l'activité. Bien
que l'allongement global de T2 soit dû à
l'effet cumulatif de toutes les compo-
santes, il doit être clair que l'on part du
principe que les allongements de T2 sont
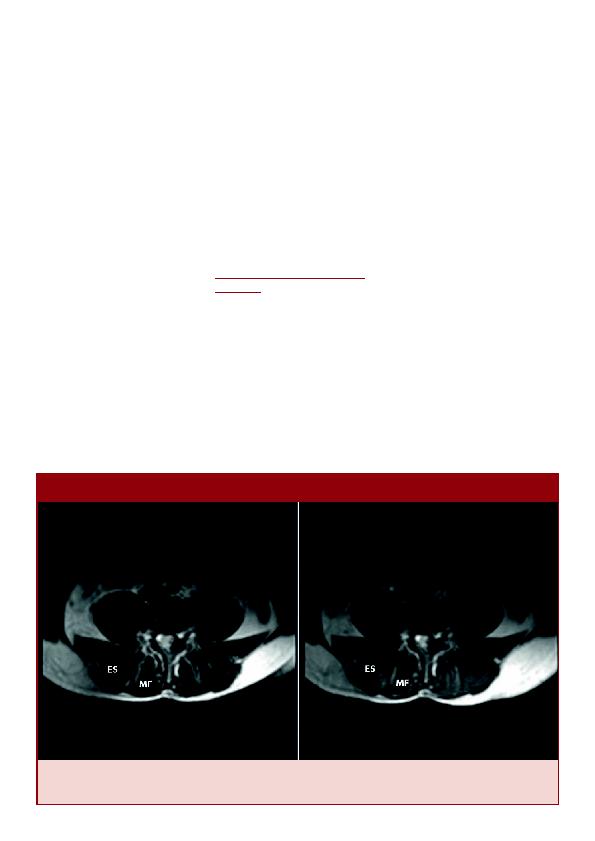
allant de 10,1 à 161,6ms; matrice 128 x 128 et FOV 256mm).
du rachis (ER). Bien que les modifications de l'intensité du signal soient subtiles, l'utilisation du calcul des valeurs T2 les
rend quantifiables.